Gulati Lab
The Gulati Lab is a systems neuroscience laboratory interested in understanding the motor basis of behavior. The lab uses cortical and spinal electrophysiology to understand the neural basis of motor control and learning, as well as sleep-related consolidation of a motor skill.
The Gulati Lab is also investigating novel neuroengineering strategies for repair and functional recovery after brain injury (stroke) and spinal cord injury that utilize development of spinal/cortical motor prostheses, brain machine interfaces and neurorobotics.

Personal Statement
We are conducting preclinical and clinical research with the overarching goal of developing neurotechnology that can improve stroke rehabilitation. Using rodent models, we investigate neural mechanisms of stroke recovery and utilize this knowledge to optimize clinical neurorehabilitation."
Tanuj Gulati, PhD
Breakthrough Research
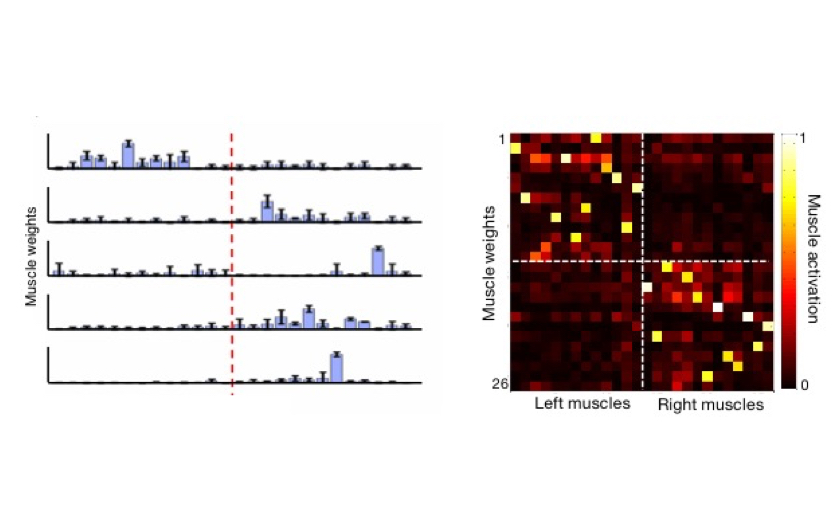
The Gulati Laboratory research interests are in motor-systems neuroscience, especially in how motor control and motor learning are enacted by motor networks in intact and injured states. The Gulati Lab is using electrophysiology tools in rodents and humans to study these processes at the single neuronal and network level.
Publications
Learn more about the recent research publications from the Gulati Lab.
Laboratory Team
Learn more about the scientists, faculty members, investigators and other healthcare professionals of the Gulati Lab, whose dedicated efforts lead to groundbreaking discoveries.
Job Opportunities
We are actively recruiting students and postdocs to be our colleagues and expand lab knowledge and expertise to make important breakthroughs.
Collaborations
The Gulati Lab collaborates with a wide range of investigators within Cedars-Sinai and institutions around the world.
Contact the Gulati Lab
127 S. San Vicente Blvd.
Advanced Health Sciences Pavilion, Eighth Floor
Los Angeles, CA 90048